
TURION OVERWINTERING OF AQUATIC CARNIVOROUS PLANTS
Carnivorous Plant Newsletter 28: 19-24 (1999)
LUBOMÍR ADAMEC
Academy of Sciences of the Czech Republic, Institute of Botany, Dukelská 135, CZ-379 82 Třeboň, Czech Republic; E-mail: adamec@butbn.cas.cz
Keywords: ecology: Aldrovanda vesiculosa, Utricularia.
Introduction
Overwintering buds (turions; Latin turio=shoot) are vegetative dormant organs produced by perennial aquatic plants. Turions are formed in subtropical to polar zones as a response to unfavorable ecological conditions, and protect fragile plant shoots from freezing and decaying. Turions are modified shoot apices. Both morphologically and physiologically, turions of free-floating aquatic plants, and especially of Utricularia and Aldrovanda, are the most modified ones of all. During the winter they separate from the mother shoot, which totally decays afterwards.
Turions of aquatic carnivorous plants are more or less spherical, sturdy organs 1-20 mm in size, and form by the extreme condensation of very short modified leaves in the shoot apex at the end of the growing season (Figure 1). Their leaves bear only rudimentary traps. Turions are frost resistant, but their main ecological function is to sink to warmer water. At the end of winter they rise to the water surface where they sprout in warmer water. They also behave as propagules and are spread by waterways or water birds.
Initiation and Ripening of Turions
In central Europe, turions formation is preceded by the production of distinctly shortened internodes in late August. The turions are usually formed in September after the growth rate of shoot apices reduces. Cooler and shorter days induce turion formation. N or P deficiency in water apparently does not have the influence on turion formation of aquatic carnivorous plants that it does on duckweeds. The relative importance of temperature decrease and short days probably differs in different species. In U. vulgaris in Canada (52°N), turions begin to form as early as August as a result of cooling waters and shortening day length.
Figure 1: Aldrovanda vesiculosa turions stored in refrigerator over winter, late April 1997.
In thermophilous Aldrovanda, low water temperature is crucial and decreasing light might be an important effect, but the effect of short days is negligible. Turions formed about three weeks later (at the end of September) in Aldrovanda grown in a relatively warm outdoor culture at Třeboň (49°N, Czech Republic) than in plants introduced to colder shallow waters in the Třeboň region (early September). At the northernmost site at Lake Ladoga in northern Russia (61.5°N), they form as early as in mid-August and ripen in early September. In October, Aldrovanda also formed normal turions in a heated greenhouse where water temperature never fell below 18°C. When grown from turions in an aquarium on a window ledge in a heated room, normal turions were also formed in March-May, i.e. under the conditions of long (and increasing) day length. These plants formed turions in late May after they were transferred outdoors to colder water. Reports from J. A. Mazrimas growing the Japanese subtropical Aldrovanda indicate turion formation to be initiated at 16°C; however, it is not clear whether this is the day, night, or average temperature. Apparently, turion formation in Aldrovanda occurs after a long-term marked relative temperature fall (e.g. from 30 to 20°C), and not after reaching some low threshold temperature.
While the formation of turions may occur at 18-20°C, full physiological turion maturation requires lower temperatures (at least 8-10°C), lessening light, and possibly also short days. Turion maturation is accompanied by the translocation of the majority of N, P, and storage organic substances (sugars) from the dying mother shoots. As turions mature, they become more compact and contain more storage substances. In the absence of low temperatures, evergreen shoots do not decay over the winter season but remain firmly connected to the turions.
The development of turions is also controlled by humic acids. In Aldrovanda, humic acids in water accelerate the formation, maturation, and sinking of turions. In spring, humic acids mildly inhibit turions from rising to the water surface to sprout (by altering their buoyancy); humic acids also enhance the growth of already germinated turions. Humic acids (and maybe also tannins) thus play an important role as exogenous phytohormones leading to optimal seasonal Aldrovanda development.
Sinking of Turions
It is interesting that turions of seven similar European aquatic carnivorous plant species have three different strategies of autumnal sinking and spring floating. In two strategies, sinking and floating are passive and are not directly controlled by turions.
a) U. australis, U. vulgaris, U. intermedia, and U. ochroleuca: in this group, ripe turions are always less dense than water and are firmly connected to the mother shoots. As the mother shoots gradually decompose, they become denser than water and drag the turions to the bottom. By early spring, the turions separate and float to the surface. The turions can reach the water surface as soon as the ice cover has melted.
b) U. minor and U. bremii: turions start forming in late August with an unusual structure in the shoot apices (Figure 2). Antler-like branched leaves form a basket (1-2 cm across) enclosing the developing turion. In late September and October, light ochre-colored mother shoots die and drag the turions to the bottom. In autumn the turions break free, but remain entangled in the apical baskets. They float to the surface only after the baskets decompose.
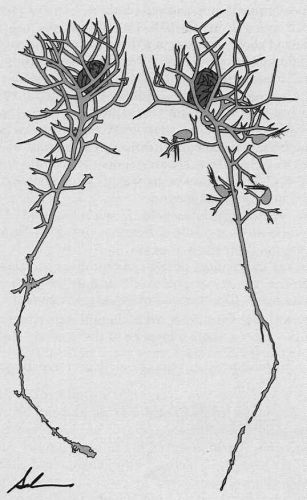
Figure 2: Utricularia bremii from outdoor culture, late October 1997. Turions are surrounded by baskets of apical leaves. Drawn from herbarium specimens by E.M. Salvia.
c) A. vesiculosa: in this species, a remarkably active way of sinking and rising has developed. In autumn ripe turions break off the dying mother shoots at the water surface. After a few days, they sink gradually to the bottom. Their high density is not only caused by their high starch content but also, presumably, by their expulsion of excess gases. In time, the turions are slightly obscured in the sediments. In April-May, they respond to the warming water, and within a few days rise to the surface.
Composition and Resistance of Turions
Turions of aquatic carnivorous plants are storage organs. While the dry weight (DW) of summer shoots is only 8-12% of the fresh weight, it is 25-46% of the fresh weight in autumn. The high proportion of DW in autumn is caused by the accumulation of starch (25-32% DW) and free sugars (glucose, fructose, sucrose; in total 7-14% DW). Stores of N are in the form of amino acids (arginine, lysine). Over winter, the metabolism of turions is very low and probably anaerobic. Their respiration rates at 4-5°C are about one order of magnitude lower than those of summer shoots at 20°C. Yet, the total content of saccharide energy sources in turions gradually decreases to 15-24% DW over winter. In this way, the DW proportion also decreases to about 18-20%. The spring starch content is rather low (about 8-11% DW) but the free-sugar content is increased (7-16% DW).
Ripe turions of aquatic carnivorous plants are frost-resistant. Ripe Aldrovanda turions do not survive storing at -10°C in water or -12°C in humid air. When they are overwintered in the wet bottom of a drained tray (our winters are moist so such exposed turions do not dessicate) or in the field where they are gradually adapted to frosts, they fully survive frosts of -10 to -15°C. Aldrovanda turions survive -4°C for long period. U. vulgaris turions exhibit an increasing resistance to -8°C in the course of winter but they do not survive -12°C (similar behavior has been observed in U. australis turions). However, their frost-resistance is very low when they are germinating. The high resistance of turions to weak frosts of -2 to -4°C could be used to store them for several years, since turions survive for only about 10-12 months when stored at 3-5°C. This suggests turions at natural sites might only survive from one season to the next one, not longer.
Turions are drought-resistant. In U. vulgaris and Aldrovanda, turion-like organs are also formed as a result of summer drought. They are not dormant, and in water, they resume their growth again.
Turions usually overwinter under unfavorable conditions of anoxia, a strongly reduced medium (i.e. low redox potential), and in the presence of toxic substances (e.g. H2S or acetic acid). Ripe turions of aquatic carnivorous plants are able to tolerate these unfavorable conditions even though experimental evidence for this is still lacking. In the spring, their tolerance of these conditions decreases.
Dormancy States and Their Hormonal Regulation
After turion formation, two dormancy states can be distinguished. They are controlled by endogenous levels of stimulating and inhibiting phytohormones. The dormancy states (and their hormonal patterns) have been described in detail for Canadian U. vulgaris. Turions enter a state of innate dormancy at the end of August, when their growth is blocked by endogenous factors in the turions (even though external ecological conditions are still favorable to growth). During innate dormancy, aging and dying of mother shoots (and their sinking to the bottom) occurred. High levels of abscisic acid (the inhibitory phytohormone) and low levels of free gibberellins, auxin, and cytokinins, occur in this dormancy state. Abscisic acid has the principal influence, as it induces turion formation. High temperatures (30°C) can temporarily break innate dormancy, but is followed by the formation of turions again. Innate dormancy was only broken by the combination of short days and low temperatures. In field collected U. vulgaris, this dormancy state was broken, as early as in late October under favorable conditions. In fact, the turion short-day requirement to break innate dormancy was fully satisfied as early as early September, whereas the low-temperature requirement was met after 55 days. In Aldrovanda turions, both in the field and outdoor culture, the short-day requirement was fully satisfied in early November. Turions stored at 4°C in darkness germinated at the end of January.
After their innate dormancy, turions of Canadian U. vulgaris enter a state of imposed dormancy at the end of October. This is maintained by low, unfavorable temperatures. This dormancy state is characterized by a decreasing level of abscisic acid and increasing levels of gibberellins, auxin, and cytokinins. When the imposed dormancy is broken by a temperature increase, the first turions germinate after two days. Breaking imposed dormancy begins germination. Low levels of abscisic acid and high levels of the other three stimulatory phytohormones correspond to this phase. In Aldrovanda, imposed dormancy breaks at 12-15°C.
Breaking imposed dormancy (which may not require light) is connected with the activation of respiration and partial starch conversion to free sugars. The density of Aldrovanda turions is reduced within 1-2 days so turions rise to the surface where they germinate in of warmer water, high light intensity, and increasing day length. In duckweed turions, a small gas bubble (presumably CO2 released from anaerobic respiration) is produced which pulls the turions to the water surface. This process can occur in darkness, but is more effective in light. The density of Aldrovanda turions may be similarly reduced by the production of CO2 into the fine air spaces in leaves, expelling water from them. A photosynthetic origin of this gas is unlikely for Aldrovanda. Moreover, the net photosynthetic rate of dormant turions of bladderworts and Aldrovanda is very low or negative, even under optimum conditions. When turions are exposed to light after breaking imposed dormancy, they resume shoot growth (Figure 3). Although turion germination in itself is generally controlled by the photoreceptor phytochrome, great differences in light requirements exist between European carnivorous plants species. Turions of U. australis and Aldrovanda germinate at higher temperatures in light only. Those of U. vulgaris, U. minor, and U. bremii start germinating also in darkness and U. vulgaris even germinates in the refrigerator at 4°C.
Turion Ecology
Recent preliminary findings indicate only a minor percentage of Aldrovanda turions to overwinter successfully in the field (Kamiński, personal communication, 1994). This may be true for other aquatic carnivorous plant species. Therefore, overwintering may be a critical phase of the growing cycle of aquatic carnivorous plants. This has also been discovered by many carnivorous plant growers. In orientation growth experiments in enclosures with intact bottoms at natural sites in Poland, only 0-20% of the Aldrovanda turions survived. In growth experiments in bottomless enclosures in the Czech Republic, 0-70% of turions (on average, approximately 20-30%) survived. Some turions in the Třeboň region (S. Bohemia) hibernated on the top of the wet bottoms and were grazed by small rodents. In the Doksy region (N. Bohemia), turions were commonly grazed by ducks. Herbivores rarely graze adult Aldrovanda but their grazing is focused on the energy-rich turions. When turions overwintered below water at the same or similar sites in the Třeboň region, their survival was much higher (35-100%).
The rising of turions of aquatic carnivorous plants from the bottom is, in some species, their most susceptible phase. Turion release from the bottom is affected by how deep they are buried. In typical habitats of aquatic carnivorous plants in shallow reed and sedge stands, the greatest part of the litter, dead leaves and stems sinks to the bottom during the winter and during elevated water levels in spring. The denser these emergent stands, the more litter is available to bury the turions and impede their rising. It also increases the anoxia at the bottom. Duckweed turions do not germinate at O2 shortage. The same may be true for carnivorous plant turions. Dense mats of filamentous algae or aquatic mosses may also impede rising turions. Aldrovanda turions entangled on the boundary between the bottom and free water may rot. However, turions of U. minor, U. intermedia, and U. ochroleuca, which often grow in very shallow waters, often germinate at the bottom.

Figure 3: Germinating turions of Aldrovanda, early May 1997.
Dense stands of reeds and sedges are not suitable habitats for summer growth of aquatic carnivorous plants because of shading, nor are they suitable for successful turion overwintering. The low overwintering rate of Aldrovanda turions was observed after the wintering site had been trodden underfoot by roe-deer. Afterwards, many of the turions germinated as late as summer.
Acknowledgments
The paper is dedicated to my colleague Dr. Jan Květ (Inst. Bot., Třeboň, Czech Republic) on the occasion of his 65th birthday and for his whole life study of wetland plant ecology and great merits in wetland protection.
References
Adamec, L. 1995, Ecophysiological Study of the Aquatic Carnivorous Plant Aldrovanda vesiculosa L., Acta Bot. Gall., 142: 681-684.
Appenroth, K.J., Hertel, W. , and Augsten, H. 1990, Photophysiology of Turion Germination in Spirodela polyrhiza (L.) Schleiden, The Cause of Germination Inhibition by Overcrowding, Biol. Plant., 32: 420-428.
Bartley, M.R., and Spence, D.H.N. 1987, Dormancy and Propagation in Helophytes and Hydrophytes, Arch. Hydrobiol. (Beih.), 27: 139-155.
Kamiński, R. 1987, Studies on the Ecology of Aldrovanda vesiculosa L. II. Organic Substances, Physical and Biotic Factors and the Growth and Development of A. vesiculosa, Ekol. Pol., 35: 591-609.
Mazrimas, J.A. 1978, Aldrovanda, Carniv Pl. Newslett., 7: 102-103.
Newton, R.J., Shelton, D.R., Disharoon, S., and Duffey, J.E. 1978, Turion Formation and Germination in Spirodela polyrhiza, Am. J. Bot., 65: 421-428.
Winston, R.D., and Gorham, P.R.1979a, Turions and Dormancy States in Utricularia vulgaris, Can. J. Bot., 57: 2740-2749.
Winston, R.D., and Gorham, P.R. 1979b, Roles of Endogenous and Exogenous Growth Regulators in Dormancy of Utricularia vulgaris, Can. J. Bot., 57: 2750-2759.
"Used with joint permission of the International Carnivorous Plant Society"
Copyright (c) 1995-2002 www.BestCarnivorousPlants.com
![]()