Photosynthetic characteristics of the aquatic carnivorous plant Aldrovanda vesiculosa
Aquatic Botany 59: 297-306 (1997)
Lubomír Adamec
Academy of Sciences of the Czech Republic. Institute of Botany, Section of Plant Ecology, Dukelská 135, CZ-379 82 Třeboň, Czech Republic
Tel.: +420 333 721156; fax: +420 333 721136; e-mail: adamec@butbn.cas.cz
Abstract
Aldrovanda vesiculosa L. (Droseraceae) is a critically endangered aquatic carnivorous plant. It is considered to be strictly stenotopic. Chemical and physical factors were measured in situ in A. vesiculosa stands at eight sites. The O2 concentration usually ranged between 0.25-0.28 mM. There was a wide range of pH of 5.04-7.60, but with a median of 7.17. Total alkalinity ranged between 0.16-4.2 meq l-1 but was mostly within 1.0-2.7 meq l-1. A high CO2 concentration was found in all stands; it ranged between 0.14-3.5 mM and was usually within 0.2-0.5 mM. A high CO2 concentration appeared to be the most important condition for rapid apical growth. It appeared to be a strict CO2 user with a CO2 compensation point of 5.9-8.2 microM, a Km of 165 + 77 microM CO2, and a maximum photosynthetic rate of 99 + 10 mmol kg-1 (FW) h-1. It was found to be photophilous and exhibited a light compensation point of 6.0 W m-2. A. vesiculosa grew in unshaded, and also shaded stands with only 18-24% of incident irradiance at the water surface. Photosynthetic rates rose from 15 to 34°C. The highest photosynthetic rate was found in young apical shoot segments and it declined toward the bases.
Keywords: Chemical and physical factors; Polish sites
1. Introduction
Aldrovanda vesiculosa L. (Droseraceae) is a critically endangered aquatic carnivorous plant, rapidly vanishing from Europe (Berta, 1961; Walters, 1979; Adamec, 1995a). It ís rootless, free-floating, and grows just below the surface in shallow standing dystrophic waters. It is considered to be strictly stenotopic (i.e., with narrow ecological amplitudes, see Walters, 1979; Studnička, 1984). It is also highly sensitive to competition with other aquatic plants that form denser stands (Studnička, 1984; Adamec, 1995a). Aldrovanda has the same life form and a similar growth strategy as the European Utricularia species. The principal feature is its fast apical growth (1-2 new whorls a day; Mazrimas, 1978) while the basal part is continuously decomposing. As adult A. vesiculosa plants are usually only 8-15 cm long (Studnička, 1984; Kamiński, 1987a) the physiological gradient (i.e., gradient in metabolic activity and nutrient content) between the growing apex and decomposing base should be very steep. The rapid apical growth of A. vesiculosa is necessary to overcome a covering with epiphytic algae and contributes to fast vegetative propagation. The European plants propagate only vegetatively by apical branching of the shoots (Berta, 1961; Adamec, 1995a). A high frequency of apical branching indicates favourable growth conditions.
Photosynthetic characteristics may predetermine and explain important ecological requirements of a plant species. In submerged plants especially, the net photosynthetic rate (PN) may be limited by a low availability of dissolved inorganic carbon (e.g., Maberly and Spence, 1983; Sand-Jensen, 1987; Pokorný and Ondok, 1991). A. vesiculosa has been characterized ecologically as a thermo- and photophilous species (Berta, 1961; Studnička, 1984; Adamec, 1995a). Water temperature in natural A. vesiculosa stands may reach 30°C during a summer day and decline by 12°C during night (Studnička, 1984). It was reported that within stands dissolved O2 concentrations ranged between 0.075-0.31 mM and pH values between 5.6-6.8 (Komiya, 1966; Studnička, 1984; Kamiński, 1987a; Akeret, 1993). A. vesiculosa is able to grow in waters differing greatly in total alkalinity (TA; TA = [HCO3-] + 2 . [CO32-] + [OH-] - [H+]) from ca. 0.5 to 5 meq l-1 (Komiya, 1966; Kamiński, 1987a; Akeret, 1993). An accumulation of partly decomposed plant litter often occurs in A. vesiculosa stands (Studnička, 1984; Kamiński, 1987a). The litter releases humic acids which were found to be essential for the growth and development of A. vesiculosa (Ashida, 1937; Kamiński, 1987a,b). The typical feature of water chemistry in A. vesiculosa stands is a very high free-CO2 concentration, due to litter decomposition (0.3-0.7 mM, Komiya, 1966; 1.9 mM Akeret, 1993). High CO2 concentrations in water are considered to be a major ecological requirement of A. vesiculosa and ensure its rapid apical growth (Adamec, 1995a).
To elucidate the ecological tolerances of A. vesiculosa, pH, TA, O2 and CO2 concentrations, irradiance in the stands, and water temperature were measured in situ in its stands at nearly all Polish sites. Basic photosynthetic characteristics of A. vesiculosa, including CO2 vs. HCO3- use, dependence of PN on CO2 concentration, irradiance and temperature responses, and shoot segment position were also determined.
2. Materials and methods
2.1. Field investigations
Ecological factors were investigated in A. vesiculosa stands in the peak of the growing season at the following sites (Adamec, 1995a): (a) Augustów-Suwalki Lake District in NE Poland (53°55'N, 23°00'E): Lakes Miklaszówek, Kruglak, Orle, and Krzywe; 11-12 July 1994; (b) Miedzychodzko-Sierakowskie Lake District in W Poland (52°33'N, 16°11'E): Lake Ostrowo, 14 July 1994; (c) Leczna-Wlodawa Lake District in E Poland (51°26'N, 23°06'E): Lakes Dlugie, Moszne and Brzeziczno, 14-16 June 1993.
Investigations were carried out in relatively dense A. vesiculosa stands (> 20 plants m-2) with various types of accompanying vegetation. The pH, dissolved [O2], and water temperature were measured 1-2 cm below the surface at between 12:00-18:00 h. Irradiance was measured with a cosine-corrected PAR sensor (Inst. Microbiol., Třeboň, Czech Republic) both above the vegetation and at the water surface, randomly at six to eight places in a stand from 12:00 to 18:00 h during clear or slightly cloudy weather. The results are expressed as percentages of incident PAR penetrating to the water surface. TA in water samples was estimated by Gran titration (Talling, 1973). CO2 concentrations were calculated from the pH and TA values after Helder ( 1988).
2.2. Plant cultivation
A. vesiculosa plants for cultivation were collected in Lake Dlugie in E Poland. Plants were cultivated outdoors in a 1-m2 plastic container. A 3-cm layer of Carex gracilis and Carex rostrata litter, over 5-cm of sand, was used as the substrate. This substrate maintained the pH of the water between 6.8-7.4, the TA 0.7-1.2 meq l-1, and the [CO2] at 0.1-0.25 mM. Water depth was kept at 20-25 cm and tap water (pH 7.6, TA 0.8 meq l-1) was added to compensate for water loss. The container was sparsely planted with C. gracilis and Phragmites australis to simulate natural conditions. The cultivated plants were 10-15 cm long, with numerous branches.
2.3. Photosynthetic measurements
Three types of photosynthetic measurements were performed: outdoor end-pH of CO2 vs. HCO3- compensation points (CP), laboratory pH-drift measurement of CO2 vs. HCO3- CP, and measurements of PN as dependent on CO2 concentration, irradiance, temperature, and shoot segment position. Healthy unbranched apical shoot segments about 2.5 cm long, with 5-6 whorls of mature leaves with fully developed traps (3-4.5 mm long), were used as standards. Dry weight (DW, 80°C) was 8.1 % of fresh weight (FW). In the segments, all traps were closed, and some contained prey.
CO2 vs. HCO3- compensation points were estimated in the light at 1 mM NaHCO3, with 0.1 mM KCl (Adamec and Ondok, 1992). Air volume of ca. 1 ml was let in the closed tubes to reduce O2 accumulation in water. After a 4-h exposure at 24 + 1°C in natural light (80-100 W m-2 of PAR), the end-pH values were measured (Allen and Spence, 1981). The CO2 CP was also estimated in duplicated pH-drift measurements when CO2-based PN was calculated from a gradual pH rise in a 40 ml closed stirred chamber in the same solution as above (for the details see Adamec, 1993). The temperature was kept at 22 + 0.1°C and photon irradiance was 120 W m-2 at the level of the plants (four apical segments).
The dependence of PN on CO2 concentration, irradiance, and temperature was measured in a 8.6 ml closed stirred chamber in the above solution as the rate of change of [O2] (e.g., Allen and Spence, 1981). One apical segment was used and the solution was renewed before each measurement. Initial [O2] was ca. 0.25 mM. Various [CO2] (10-1000 microM) in the solution were prepared by bubbling with CO2 to a calculated pH value (Helder, 1988). Each shoot segment was exposed successively to increasing [CO2]. Various photon irradiances were produced using a neutral light filter and changing the distance between the light source and the chamber. The influence of temperature on PN and dark respiration (DR) was investigated between 15-34°C. Starting with the lowest temperature, each shoot segment was exposed first to darkness and then to light. Generally, 22.0 + 0.1°C, 120 W m-2, and 0.25 mM CO2 were applied as standards. Measurement of PN in some apical shoot segments was followed by PN measurement in excised 6-mm long apices and isolated traps from the same apical segments and, also, in older shoot segments with the seventh to ninth and 10th-12th whorls of leaves from the same shoots. For 15-20 min before each measurement, shoot segments were exposed in the solution with a low CO2 concentration (pH ca. 8.0) to the same irradiance as during the next measurement.
All measurements were undertaken in quadruplicate and rates are expressed on a fresh weight basis. The plot of PN against CO2 concentration and irradiance was fitted by a nonlinear regression (ENZFITTER programme) according to Platt et al. (1980). The Michaelis-Menten model, modified to include dark respiration, was used to fit the data PN vs. CO2 concentration, while the model of exponential light saturation with three parameters and dark respiration was used to fit the data PN vs. irradiance. Since the latter model does not allow the calculation of Pmax (maximum PN) this parameter was calculated using the hyperbolic model with two parameters and DR.
3. Results
The chemical and physical environment varied considerably both within A. vesiculosa stands at one site and between all sites (Table 1). Oxygen concentration was usually between 0.25-0.28 mM but in Lakes Moszne and Brzeziczno, it was below 0.09 mM. The lakes in the Leczna-Wlodawa Lake District (Lakes Dlugie, Moszne, Brzeziczno) differed in pH, TA, and [CO2 ] from those at the other sites. The former group was characterized by low pH (5.0-6.4) and TA (0.16-1.01 meq l-1) and extremely high [CO2] (1.09-3.50 mM). At the other sites, the pH values (7.0-7.6) and [CO2] (0.14-0.62 mM) in various stands were rather similar, although the TA values sometimes differed (1.55-4.23 meq l-1). Although A. vesiculosa often grew in unshaded water, it also grew where incident PAR was only 18-24% due to shading from emergent vegetation (Table 1).
Table 1. Chemical and physical factors in A. vesiculosa stands in Polish lakes
|
Lake |
n |
[O2] (mM) |
pH |
Total alkalinity (meq l-1) |
[CO2] (mM) |
PAR (%) |
|
Miklaszówek |
8 |
0.30 (0.24-0.34) |
7.33 (7.19-7.60) |
4.23 |
0.44 (0.24-0.62) |
24 (3) 18-28 |
|
Kruglak |
4 |
0.28 (0.18-0.39) |
7.25 (7.14-7.46) |
2.67 |
0.35 (0.22-0.48) |
36 (3) 24-49 |
|
Orle |
1 |
0.27 |
7.30 |
2.42 |
0.27 |
n.d. |
|
Krzywe |
1 |
0.26 |
7.59 |
2.50 |
0.14 |
n.d. |
|
Ostrowo |
5 |
0.29 (0.26-0.34) |
7.10 (7.00-7.28) |
1.60 |
0.28 (0.18-0.36) |
33 (1) |
|
Dlugie |
3 |
0.26 (0.23-0.30) |
5.93 (5.80-6.40) |
1.01 |
2.55 (1.09-3.35) |
n.d. |
|
Moszne |
2 |
0.09 (0.022-0.15) |
5.69 (5.68-5.70) |
0.57 |
2.80 (2.44-3.16) |
n.d. |
|
Brzeziczno |
1 |
0.08 |
5.04 |
0.16 |
3.50 |
n.d. |
n = number of investigated stands; PAR = % of incident PAR penetrating to the water surface; [CO2] = CO2 concentration calculated from pH and total alkalinity; n.d. = not determined. Mean values and ranges of values are shown. For PAR, the number of measurements are shown in parentheses.
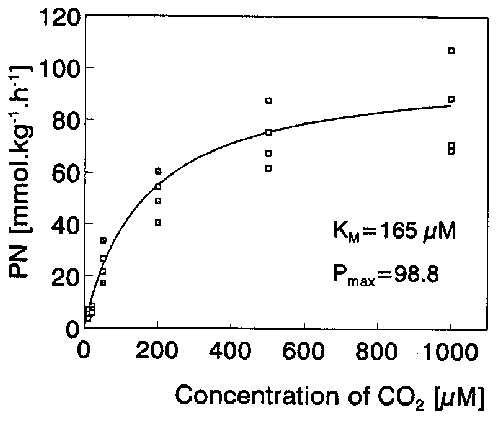
Fig. 1. Photosynthetic rates (FW-based) in A. vesiculosa apical segments as dependent on CO2 concentration at 120 W m-2 of PAR and 22°C. Km, half-saturation constant; Pmax, calculated maximum rate.
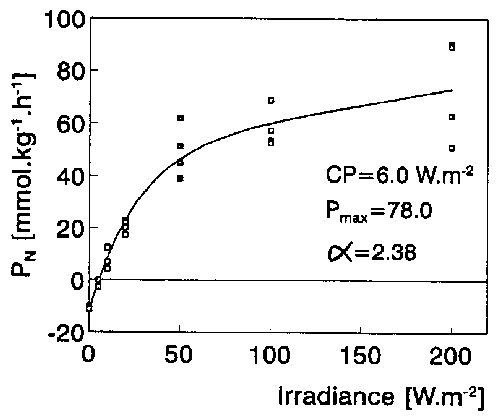
Fig. 2. Photosynthetic rates (FW-based) in A. vesiculosa apical segments as dependent on irradiance of PAR at 0.25 mM CO2 and 22°C. CP, light compensation point of photosynthesis; α, the initial slope of the curve. Pmax, was calculated using a different model (see Section 2.3).
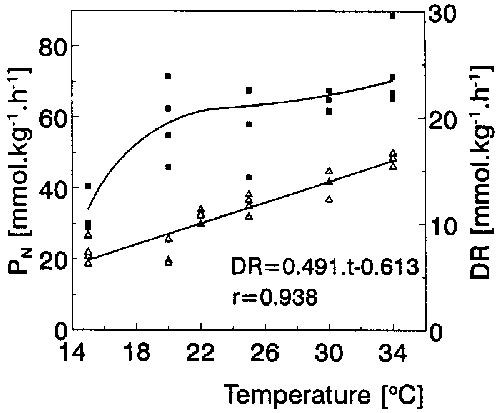
Fig. 3. Photosynthetic (squares) and dark respiration (DR, triangles) rates (FW-based) in A. vesiculosa apical segments as dependent on temperature at 120 W m-2 of PAR and 0.25 mM CO2. r is the correlation coefficient of linear regression of DR. The PN curve was fitted empirically by a polynomial function.
The same end-pH values (8.45-8.58) and CO2 compensation points (5.9-8.0 microM) were found in both laboratory pH-drift experiments and outdoor end-pH measurements. The small difference in CO2 CP between the Polish (7.2 + 0.9 microM) and Italian plants (5.9 + 0.8 microM) was not statistically significant at p < 0.05. These results suggest that A. vesiculosa is a CO2-only user. The dependence of PN on CO2 concentration was modelled using the Michaelis-Menten equation (Fig. 1). The half-saturation constant Km was 165 + 77 (SEM) microM CO2 and Pmax 98.8 ± 9.5 (SEM) mmol kg-1 (FW) h-1. PN was not apparently inhibited at the high CO2 concentration of 1 mM. A. vesiculosa was found to be markedly photophilous (Fig. 2): its light CP was 6.0 W m-2 and PN was not light-inhibited at 200 W m-2 of PAR. Pmax of the light curve was 78.0 + 5.1 (SEM) mmol kg-1 h-1 and the slope (α) of the initial linear part was 2.38 + 0.56 (SEM) mmol kg-1 h-1. Its PN was rising within the range of 15-34°C (Fig. 3). As opposed to linear dependence of DR on temperature (DR = 0.491 t - 0.613; r = 0.938), the temperature curve of PN was logarithmic with a conspicuous threshold between 15-20°C (Q10 = 3.3) whereas Q10 was only 1.1 between 20-30°C. The Q10 value of DR was 1.8 between 20-30°C. Of all investigated shoot segments, the highest photosynthetic rate (on the FW unit) was found in the standard of 2.5 cm apical segments (Table 2). The PN of excised apices as well as of excised traps were only 67% of that in the whole apical segments. The PN also decreased moderately in older shoot segments.
Table 2. A comparison of photosynthetic rate in successive shoot segments and traps of A. vesiculosa at 22°C, 120 W m-2, and 0.25 mM CO2. PN of the standard apical segments (69 mmol kg-1 h-1) was taken as 100%
|
Shoot segment |
PN (%) |
|
Apices ca. 6 mm long |
66.8 |
|
Apices with six whorls of leaves |
100 |
|
Seventh to ninth whorls |
85.2 |
|
10th-12th whorls |
75.5 |
|
Traps of first-sixth whorls of leaves |
67.0 |
4. Discussion
A. vesiculosa in natural stands can grow within a very wide range of pH, TA, [O2] and [CO2], in contrast with its common ecological characteristic as a stenotopic species (Studnička, 1984). After the chemical factors, all the sites may be subdivided into two distinct groups: Lakes Dlugie, Moszne, Brzeziczno, and the other lakes in northeastern and western Poland. The former group of A. vesiculosa sites is typified by an extremely deep sediment (10-12 m) of partly decomposed litter. Populations in the latter group are considered as more stable (Adamec, 1995a; Kamiński et al., 1996). It is possible to conclude that pH and TA individually are not important for A. vesiculosa's occurrence and growth as has been previously reported (e.g., Mazrimas, 1978; Studnička, 1984). Yet, pH values between 6.3-6.6 were usually measured by some authors in A. vesiculosa stands (Komiya, 1966; Studnička, 1984; Kamiński, 1987a; Akeret, 1993). In laboratory in a mineral medium, the best growth of its shoots and the highest frequency of branching were found at a low pH of 3.5-5.5 (Kamiński, 1987a). However, its most abundant populations (Lakes Kruglak, Miklaszówek, Ostrowo, and Dlugie) grew at the pH between 5.8-7.5 and TA 1-4 meq l-1 (Table 1).
High [CO2] seem to be crucial for A. vesiculosa. In its stands, they never fell below 0.14 mM, were usually between 0.2-0.5 mM, and sometimes reached 3.5 mM (Table 1; cf. Roelofs, 1983). It is likely that such high [CO2] in the stands were caused by deep sediments consisting of litter. The occurrence of A. vesiculosa in all sites was confined to spots with a high [CO2]. Plants were totally absent from the open lake water which had a distinctly higher pH and lower [CO2] (Lake Miklaszówek: pH 7.77, CO2 0.10 mM; Lake Orle: pH 7.83, CO2 0.08 mM; cf. Table 1). On the exact boundary of its occurrence in Lake Ostrowo, A. vesiculosa was growing in a dense mat of Spirogyra sp., the pH (8.2-8.9) and [CO2] (0.004-0.023 mM) of which represent presumably ecological limits for its survival.
The data on shading shown in Table 1 represent only an estimate of the upper limit of shading for vigorous growth of A. vesiculosa. In densest stands of A. vesiculosa in Lakes Miklaszówek and Kruglak (128 and 50 plants m-2, respectively), the level of 18-24% of incident PAR indicates probably a minimum light requirement. Thus, A. vesiculosa cannot be characterized as a strict heliophyte (Studnička, 1984) and its tolerated level of shading lies between 20-100% of incident PAR. The low values correspond to levels of shading in loose reed or Carex sp. stands, which are considered its typical and most favourable habitat (e.g. Afanasyev, 1953; Berta, 1961; Fiirst, 1986; Kamiński, 1987a,b). However, overgrowing of lake littorals by dense stands of helophytes due to accelerated lake succession probably has caused the decline of A. vesiculosa in many lakes (Adamec, 1995a). In the A. vesiculosa stands investigated, the mean surface water temperature was 26.8°C (24.2-28.9°C) during the warmest summer days. Mean water depth was 0.21 m (0.05-0.5 m). The waters where A. vesiculosa grows may be characterized as oligo- to mesotrophic (Komiya, 1966; Studnička, 1984; Kamiński, 1987a; Akeret, 1993). Since carnivory enhanced the growth of A. vesiculosa considerably (Kamiński, 1987b), it is possible to consider that the quantity of prey is more important for its vigorous growth than the level of main nutrients in water.
The values of CO2 CP (5.9-8.0 microM) of A. vesiculosa lie within the range found for strict CO2 users among aquatic plants (Maberly and Spence, 1983). Ecologically related aquatic Utricularia species exhibit the same or lower CO2 CP (1.5-6.7 microM in Utricularia purpurea, Moeller, 1978; 1.8-3.3 microM in Utricularia australis, 2.3-4.5 microM in Utricularia minor, and 7.2 microM in Utricularia vulgaris, from Adamec, 1995b). In A. vesiculosa, the Km for CO2 uptake (0.17 mM) and Pmax (99 mmol kg-1 h-1) are comparable with the data in other aquatic plants (Km 0.05-0.30 mM; Pmax 30-110 mmol kg-1 (FW) h-1 (Sand-Jensen and Gordon, 1986; Madsen, 1991; Pokorný and Ondok, 1991). Its photosynthetic quotient (O2/CO2) of about 0.76 corresponds to published values (Pokorný et al., 1989). Unambiguously, A. vesiculosa was found to be markedly photophilous (Fig. 2). Its light CP of 6.0 W m-2 is greater than for other submerged plants (between 0.3-5.1, mean 1-2 W m-2; Pokorný and Ondok, 1991).
As follows from the results, photosynthesis of A. vesiculosa is adapted to high temperatures of about 34°C. The PN decline between 15-20°C (Q10 = 3.3) could be related to the minimum temperature limit of its growth (ca. 16°C), since turion formation starts below this limit (Mazrimas, 1978). In most submerged species, the maximum PN is reached between 20-30°C (Pokorný and Ondok, 1991). In A. vesiculosa, both the apices and traps of the apical segments reached only 67% of PN of the whole segments (Table 2). As the ratio of traps to the total FW was 45.8% the contribution of the traps to the total PN was only 30.7%. Similarly in Utricularia macrorhiza (syn. U. vulgaris), the PN of bladders was 60-70% of young leaves and of apices only 35-60% (Knight, 1992). Friday (1989) confirmed that bladders in U. vulgaris age faster than leaves.
As shown in this study, A. vesiculosa is a strict CO2 user and its photosynthetic capacity is relatively high under favourable conditions of high CO2 concentration, irradiance, and temperature. Obviously, the high PN is a necessary condition for its very fast apical growth of 1-2 whorls a day (Mazrimas, 1978; cf. Friday, 1989). Otherwise a thorough reutilization of carbohydrates (and also mineral nutrients) must take place from the aged shoot bases. It follows from the photosynthetic characteristics of A. vesiculosa (Figs. 1-3) that with a reduction of the ambient CO2 concentration below about 0.15 mM, the irradiance below about 40-50 W m-2, and the temperature below about 20°C will lead to a steep decrease in its PN. This finding corresponds to the ecological relations found in natural A. vesiculosa stands (cf. Table 1). When the mean ecological stand factors (0.3-0.4 mM CO2, 30-40% of penetrating PAR, 24-26°C) are introduced to its photosynthetic relations the stand PN of about 59-68 mmol O2 kg-1 (FW) h-1 can be assessed. It is about 60-69% of the Pmax (for CO2), a very high value as compared to stands of other submerged plants (cf. Pokorný and Ondok, 1991). A model calculation can be made to show whether an adult plant consisting of the apex and 12 whorls of leaves (ca. 170 mg FW) is able to produce two new whorls (ca. 26 mg FW = 2.1 mg DW = 0.80 mg C; carbon = 38% DW, Moeller, 1978) a day, on the basis of the PN of 59 mmol O2 kg-1 h-1, DR 10.9 mmol kg-1 h-1, 12-h light: l2-h dark period, photosynthetic quotient 0.76, and the rates from the Table 3. Thus, the plant can theoretically fix about 1.46 mg C (122 micromol CO2 ) daily, more than its daily maximum requirement of 0.8 mg C, even when no reutilization occurs.
Great losses of A. vesiculosa turions occur in its stands over winter (Kamiński and Adamec, unpublished) and their loss must be compensated for by very fast growth and propagation during the growing season. High CO2 concentration, found in A. vesiculosa stands, is considered to be the most important factor contributing to its very fast growth and propagation. Several aquatic carnivorous species appear to be CO2-only users although they occasionally grow in moderately alkaline waters with low CO2 concentration (Adamec, 1995b). Under such conditions, carnivory may also be means for obtaining additional carbon.
Acknowledgements
The author is very grateful to Dr. R. Kamiński (Wroclaw, Poland) for organizing the field excursions and providing some unpublished data and to Mr. C. Breckpot (Haaltert, Belgium) for technical assistance and providing some literature. Sincere thanks are due to Dr. Naomi Rea (Darwin, NT, Australia) for correction of English. Thanks are also due to Dr. L. Pechar (Třeboň) for statistical treatment of the results. The author is also grateful to Prof. G. Bowes (Gainesville, FL, USA), Dr. S.C. Maberly (Ambleside, UK), and another reviewer for valuable comments on the manuscript. This research was funded by the Academy of Sciences of the Czech Republic (project no. 605401).
References
Adamec, L., 1993. Rapid inhibition of HCO3- use by high concentration of free CO2 in Elodea canadensis. Aquat. Bot. 45, 311-324.
Adamec, L., 1995a. Ecological requirements and recent European distribution of the aquatic carnivorous plant Aldrovanda vesiculosa L.-A review. Folia Geobot. Phytotax. (Prague) 30, 53-61.
Adamec, L., 1995b. Photosynthetic inorganic carbon use by aquatic carnivorous plants. Carniv. Plant Newslett. (Fullerton, USA) 24, 50-53.
Adamec, L., Ondok, J.P., 1992. Water alkalization due to photosynthesis of aquatic plants: The dependence on total alkalinity. Aquat. Bot. 43, 93-98.
Afanasyev, K.S., 1953. New site of Aldrovanda vesiculosa L. in USSR. Bot. Zh. (Leningrad) 38, 432-434, In Russian.
Akeret, B., 1993. Ein never Fundort von Aldrovanda vesiculosa L. in der Nordschweiz and einige Bemerkungen zu Stratiotes aloides L. Bot. Helv. 103, 193-199.
Allen, E.D., Spence, D.H.N., 1981. The differential ability of aquatic plants to utilize the inorganic carbon supply in fresh waters. New Phytol. 87, 269-283.
Ashida, J., 1937. Studies on the leaf movement of Aldrovanda vesiculosa L.: III. Reaction time in relation to temperature. Bot. Mag. (Tokyo) 51, 505-513.
Berta, J., 1961. Beitrag zur Őkologie and Verbreitung von Aldrovanda vesiculosa L. Biológia (Bratislava) 16, 561-573.
Friday, L.E., 1989. Rapid turnover of traps in Utricularia vulgaris L. Oecologia 80, 272-277. Fürst, R., 1986. Aldrovanda vesiculosa. Taublatt 5, 14-15.
Helder, R.J., 1988. A quantitative approach to the inorganic carbon system in aqueous media used in biological research: Dilute solutions isolated from the atmosphere. Plant Cell Environ. 11, 211-230.
Kamiński, R., 1987a. Studies on the ecology of Aldrovanda vesiculosa L.: I. Ecological differentiation of A. vesiculosa population under the influence of chemical factors in the habitat. Ekol. Pol. 35, 559-590.
Kamiński, R., 1987b. Studies on the ecology of Aldrovanda vesiculosa L.: II. Organic substances, physical and biotic factors and the growth and development of A. vesiculosa. Ekol. Pol. 35, 591-609.
Kamiński, R., Adamec, L., Breckpot, C., 1996. Report on recent sites of Aldrovanda vesiculosa L. in Poland. Fragm. Flor. Geobot. (Kraków) 41, 291-294.
Knight, S.E., 1992. Costs of carnivory in the common bladderwort, Utricularia macrorhiza. Oecologia 89, 348-355.
Komiya, S., 1966. A report on the natural habitat of Aldrovanda vesiculosa found in Hanyu City. Amatores Herb. (Kobe) 27, 5-13.
Maberly, S.C., Spence, D.H.N., 1983. Photosynthetic inorganic carbon use by freshwater plants. J. Ecol. 71, 705-724.
Madsen, T.V., 1991. Inorganic carbon uptake kinetics of the stream macrophyte Callitriche cophocarpa Sendt. Aquat. Bot. 40, 321-332.
Mazrimas, J.A., 1978. Aldrovanda. Carniv. Plant Newslett. (Fullerton, USA) 7, 102-103.
Moeller, R.E., 1978. Carbon uptake by the submerged hydrophyte Utricularia purpurea. Aquat. Bot. 5, 209-216.
Platt, T., Gallegos, C.L., Harrison, W.G., 1980. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 38, 687-701.
Pokorný, J., Ondok, J.P., 1991. Macrophyte Photosynthesis and Aquatic Environment. Academia, Prague, p. 120.
Pokorný, J., Orr, P.T., Ondok, J.P., Denny, P., 1989. Photosynthetic quotients of some aquatic macrophyte species. Photosynthetica 23, 494-506.
Roelofs, J.G.M., 1983. Impact of acidification and eutrophication on macrophyte communities in soft waters in The Netherlands: I. Field observation. Aquat. Bot. 17, 139-155.
Sand-Jensen, K., 1987. Environmental control of bicarbonate use among freshwater and marine macrophytes. In: Crawford, R.M.M. (Ed.), Plant Life in Aquatic and Amphibious Habitats. Blackwell, Oxford, pp. 99- 112.
Sand-Jensen, K., Gordon, D.M., 1986. Variable HCO3- affinity of Elodea canadensis Michaux in response to different HCO3- and CO2 concentrations during growth. Oecologia 70, 426-432.
Studnička, M., 1984. The carnivorous plant Aldrovanda vesiculosa. Zhiva (Prague) 32, 173-176, In Czech.
Talling, J.F., 1973. The application of some electrochemical methods to the measurement of photosynthesis and respiration in fresh waters. Freshwater Biol. 3, 214-243.
Walters, S.M., 1979. Conservation of the European flora: Aldrovanda vesiculosa L., a documented case-history of a threatened species. In: Hedberg, I. (Ed.), Systematic Botany, Plant Utilization and Biosphere Conservation. Almqvist & Wiksell, Stockholm, pp. 76-82.
Copyright (c) 1995-2002 www.BestCarnivorousPlants.com
![]()